Zipf’s law refers to the phenomenon that many data sets in social and exact sciences are observed to obey a power law of the form  with the exponent
with the exponent  approximately equal to 2. In the present note I want to set out a simple Yule-Simon process (similar to one first discussed in Simon, H, 1955, On a class of skew distribution functions, Biometrika, 44:425-40) which shows clearly how Zipf’s law can emerge from urn-type processes following a similar pattern.
approximately equal to 2. In the present note I want to set out a simple Yule-Simon process (similar to one first discussed in Simon, H, 1955, On a class of skew distribution functions, Biometrika, 44:425-40) which shows clearly how Zipf’s law can emerge from urn-type processes following a similar pattern.
The simple process discussed here involves the appearance of new species within families of closely related species called genera. New species appear within genera (through evolutionary processes) which usually remain quite close in their main characteristics to the pre-existing species. However, every so often, a new species will appear which is sufficiently different from all pre-existing ones to enable it to be regarded as having started a completely new genus. We can construct a simple Yule-Simon process as a stylised version of this. Suppose that species appear one at a time and that when the number of species reaches  , the next new species will start a new genus. Therefore, when the first new genus appears, there are
, the next new species will start a new genus. Therefore, when the first new genus appears, there are  species in total. Species continue to appear one at a time, and when the number of species reaches
species in total. Species continue to appear one at a time, and when the number of species reaches  (i.e., another species have appeared), the next new species will again start a new genus. Thus, when the second new genus appears, there are
(i.e., another species have appeared), the next new species will again start a new genus. Thus, when the second new genus appears, there are  species in total. We assume this process continues indefinitely, so that when the
species in total. We assume this process continues indefinitely, so that when the 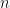 -th new genus appears, there are
-th new genus appears, there are  species in total.
species in total.
We further assume that between each genus and the next, the new species that appear will be distributed among the already existing genera in proportion to the number of species they already have (this gives rise to the characteristic feature of Zipf’s law when applied to wealth distribution in economics that `the rich get richer’). So at stage , the next species that appears will appear in the 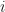 -th genus with probability
-th genus with probability

where  is the number of species already in , and
is the number of species already in , and  is simply the total number of species at stage , which is . There are opportunities for this to happen, so genus gains a new species with probability
is simply the total number of species at stage , which is . There are opportunities for this to happen, so genus gains a new species with probability

Let  denote the fraction of genera that have
denote the fraction of genera that have 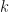 species when the total number of genera is . Then
species when the total number of genera is . Then  is the number of genera that have species when the total number of genera is , and the expected number of genera of size that gain a new species in this interval is
is the number of genera that have species when the total number of genera is , and the expected number of genera of size that gain a new species in this interval is

Now, when these genera gain the new species, they will move out of the class of genera with species, and into the class of genera with  species, so the number of genera with species will fall by
species, so the number of genera with species will fall by  . Analogously, the expected number of genera with
. Analogously, the expected number of genera with  species that will gain a new species is
species that will gain a new species is

When these genera gain a new species, they will move into the class of genera with species, so the number of genera with species will rise by . Therefore we can write a master equation for the new number  of genera with
of genera with  species at stage
species at stage 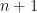 thus:
thus:
![(n+1)p_{k,n+1} = np_{k,n} + \frac{m}{m+1}[(k-1)p_{k-1,n} - kp_{k,n}]](https://s0.wp.com/latex.php?latex=%28n%2B1%29p_%7Bk%2Cn%2B1%7D+%3D+np_%7Bk%2Cn%7D+%2B+%5Cfrac%7Bm%7D%7Bm%2B1%7D%5B%28k-1%29p_%7Bk-1%2Cn%7D+-+kp_%7Bk%2Cn%7D%5D&bg=ffffff&fg=111111&s=0&c=20201002)
However, this master equation does not hold for genera of size 1. Instead, these genera obey

The second term on the right-hand side is 1 because, by definition, exactly one new genus appears at each step of the process, so there is only one entrant from the class of genera with zero species into the class of genera with one species.
We assume there is a steady state as  , in which case we get the steady state equation for
, in which case we get the steady state equation for
![(n+1)p_{k} = np_{k} + \frac{m}{m+1}[(k-1)p_{k-1} - kp_{k}]](https://s0.wp.com/latex.php?latex=%28n%2B1%29p_%7Bk%7D+%3D+np_%7Bk%7D+%2B+%5Cfrac%7Bm%7D%7Bm%2B1%7D%5B%28k-1%29p_%7Bk-1%7D+-+kp_%7Bk%7D%5D&bg=ffffff&fg=111111&s=0&c=20201002)


and the steady state equation for 


But using the steady state equation for  above we observe that
above we observe that

and substituting this back into the steady state equation for above we get

Continuing the iteration in this way we get

and using the steady state expression for 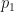 in this we get
in this we get

Since  with
with  , we can write this as
, we can write this as

Now, the gamma function is defined as

and the beta function  for
for  and
and  is defined as
is defined as

It is not too difficult to show that the two are related by the equation

and furthermore, for large  we have
we have

Comparing with the final expression for above in terms of the gamma function we see that

So we get a power law when the genera size is large, and we get Zipf’s law when the number of new entrant species is large.